
.png)


How are calcium oscillations generated and maintained
My research interrogates the mechanisms which elicit and maintain hormone-induced calcium oscillations in primary hepatocytes. In liver hepatocytes hormonal regulation of calcium signaling underlies many of the primary functions of the organ including glucose metabolism, mitochondrial physiology and gene expression. Hormones which couple to phospholipase C (PLC), Gq-linked G-protein coupled receptors (GPCRs), hydrolysis the membrane phosphoinositide lipid PIP2 to generate inositol 1,4,5-trisphosphate (IP3), and diacylglycerol (DAG), which in turn regulate cell function via calcium release and PKC activation. Hepatocytes display stimulus-strength regulated calcium oscillations and waves importantly the pattern of intracellular calcium fluctuations encodes complex spatial and temporal information to regulate cell function. We utilize single cell digital fluorescent imaging techniques combined with biochemical techniques to dissect the mechanisms which generate, maintain and regulate calcium oscillations. These studies address the ongoing debate, whether calcium oscillations arise due to calcium feedback on the IP3 receptor or as a consequence of fluctuations in IP3, hence phospholipase C activity. Our studies have provided compelling evidence that calcium oscillations in hepatocytes depend on IP3 fluctuations arising from positive calcium feedback on PLC.


The role of calcium dysregulation in the development of liver disease
Non-alcoholic fatty liver disease (NAFLD) is linked to type 2 diabetes and a number of other major comorbidities. In the liver the normal balance of carbohydrate and lipid metabolism is maintained by lipogenic hormones including insulin, and glucogenic and lipolytic hormones including glucagon and catecholamines. A common signaling pathway involved in the control of catabolic metabolism in liver involves activation of PLC by GPCRs to produces the calcium-mobilizing second messenger IP3, and DAG. These GPCRs generate cytosolic calcium oscillations in single hepatocytes, which are integrated into intercellular calcium waves that propagate across entire lobules in the intact liver. Our data show that short-term high fat diet (HFD) feeding of mice attenuates PLC-linked calcium signaling in hepatocytes and impairs propagation of calcium waves in intact liver. Zonation of liver metabolism across the porto-central axis is an essential feature of healthy liver physiology. I investigate the effects of this loss of signal transduction throughout the liver and how this effects lipid and carbohydrate metabolism and the leads to steatosis and NALFD.

.png)
Steatosis and dysregulation of metabolic zonation

A key feature of hepatic function is the segregation of different pathways across the liver lobule. Gluconeogenesis and oxidative metabolism occur primarily in the periportal zone, and glycolytic and lipogenic metabolism predominate in the pericentral zone. Our data using intravial imaging of ex vivo perfused livers, and stratification of isolated hepatocytes with cell surface zonal markers show that HFD selectively suppresses the sensitivity of pericentral hepatocytes to NE stimulation. We are investigating the effects of HFD on PLC-linked calcium signaling and its regulation of metabolic processes in zonally stratified isolated hepatocytes, as well as in the intact perfused liver. These studies will characterize the zonal organization of calcium signaling and determine how it is modified during the development of diet-induced obesity.
.png)
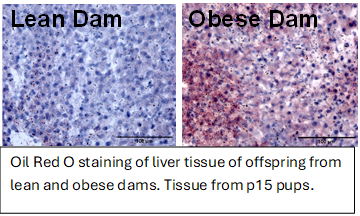
The effect of maternal obesity on liver development and function.
Developmental origins of health and disease (DOHaD)

Maternal obesity is now recognized as an independent risk factor for the development of metabolic disease in offspring. Intrauterine exposure to nutrient excess contributes to the alarming risk in pediatric obesity and fatty liver disease. The etiology of pediatric and adult metabolic associated fatty liver disease (MAFLD) differs. In adult’s steatosis and fibrosis develop primarily in the pericentral zone whilst in pediatric patients’ liver damage occurs in the periportal region. The mechanisms that drive these differences are not well understood and increasing evidence suggests the onset of pediatric MAFLD may have fetal origins. Our goal is to determine how maternal obesity affects the development of hepatic zonation using a mouse model, integrating spatial transcriptomics and metabolomics with histology and immunohistology profiling. We aim to identify non parenchymal cell populations affected by maternal obesity, determine how cell-cell communication is dysregulated, and establish how this perturbs normal hepatic zonation and metabolic function.
Calcium signaling in parasites
Plasmodia, the etiological agent of malaria, evade the human endocrine system partly by using host melatonin surges to synchronize their intraerythrocytic life cycle. The ability of melatonin to elicit calcium release in was established by Collaborators however, the molecular mechanisms of calcium release have remained elusive partly due to the lack of genetic evidence of intracellular calcium release channels. My work demonstrated that melatonin elevates inositol phosphate turnover in P. falciparum intraerythrocytic cultures. Furthermore, using photorelease of caged IP3 in intact parasites I showed equivocally IP3 releases calcium from a thapsigargin sensitive store. I have also evidenced caged IP3 dependent calcium release in Toxoplasma gondii and Trypanosoma brucei indicating a canonical IP3-dependent signaling pathway is conserved in protist parasites. Sequence homology studies have not identified IP3/Ryanodine like channels, cell surface PLC-linked receptors or G-proteins in any malaria species. However, this highlights the therapeutic potential of targeting these divergent calcium signaling molecules. I am currently investigating a number of small molecules as melatonin receptor ligands with the potential to perturb calcium homeostasis and cell cycle progression.